

论文:短须裂腹鱼产卵行为观察
短须裂腹鱼产卵行为观察
颜文斌1,2,朱挺兵2,吴兴兵2,杨德国2,陈 亮1,2
(1.上海海洋大学水产与生命学院,上海 201306;2.中国水产科学研究院长江水产研究所,农业部淡水生物多样性保护重点实验室,武汉 430223)
2014-2016年两个繁殖期,采用定点原位水下视频观察法,在四川省攀枝花市国胜河开展了短须裂腹鱼(Schizothoraxwangchiachii)产卵行为研究。结果显示,短须裂腹鱼繁殖期主要的产卵行为包括探索环境、领域争夺与防卫、配偶选择、交配、亲体护卫等。基于观察结果提出了短须裂腹鱼的产卵行为模式。
短须裂腹鱼(Schizothoraxwangchiachii);产卵行为;行为模式;水下视频
短须裂腹鱼(Schizothoraxwangchiachii)隶属于鲤科裂腹鱼亚科,主要分布于金沙江、大渡河及乌江等水系[1,2],是当地重要的经济鱼类。受过度捕捞和水电开发等因素的影响,金沙江水系短须裂腹鱼资源量急剧下降[3]。为恢复其自然种群资源,开展短须裂腹鱼增殖放流、栖息地修复等保护技术研究是目前急需开展的工作。例如,刘跃天等[4]和李光华等[5]在短须裂腹鱼的人工繁殖技术方面已经取得了一定的成果。行为学对于指导珍稀濒危鱼类的保护工作也具有重要作用,但目前对于短须裂腹鱼的行为学研究还有待深入。
鱼类产卵行为包括鱼类进入产卵场至后代孵出过程中亲鱼表现的全部行为,通常以交配行为为界,分为产前行为、交配行为和产后行为。近年来,行为学在鱼类保护实践中的应用正受到越来越多的重视。国内学者在人工环境中对唐鱼(Tanichtysalbonubes)[6]、叉尾斗鱼(Macropodusopercuiaris)[7]等观赏性小型鱼类的交配行为进行了观察描述。同时国外鱼类繁殖行为研究的对象则相对较广,既包括经济鱼类如鲑亚科[8]、银鲑(Oncorhynchuskisutch)[9]、大鳞大马哈鱼(Oncorhynchustshawytscha)[10]等,也涵盖了丽鱼(Laetacaraaraguaiae)[11]、欧洲鳑鲏(Rhodeusamarus)[12]、纹波石斑鱼(Epinephelusongus)[13]等淡水或近海常见鱼类。
动物行为与物种保护之间具有密切的关系,但在濒危物种保护实践中动物行为往往被忽视[14]。目前关于裂腹鱼类的产卵行为报道不多,而在野外进行水下视频观察的还未有报道。本研究采用野外水下视频观察的方式,在自然河流中观察短须裂腹鱼的产卵行为,以丰富短须裂腹鱼产卵行为资料,积累繁殖生物学数据,也为其野生群体保护方法及人工繁育技术改进等保护实践提供参考。
1 材料与方法
1.1 材料鱼与试验场所
选择四川省攀枝花市国胜乡某裂腹鱼类养殖基地驯养的短须裂腹鱼作为材料鱼。该批次材料鱼捕捞自雅砻江上游支流国胜河,并利用国胜河部分河道以及近岸石滩改造而成的流水池塘进行人工驯养,因此材料鱼的生长环境与自然溪流非常接近。2015年该批次材料鱼为6~8龄,均已到达成熟年龄。
行为观察地点位于国胜河热水塘村段(27°06′49.34″N;101°29′55.06″E;海拔1 495 m)。该河段水质优良,除降雨天气外大部分时间清澈见底。行为观察试验期间(2015年12月—2016年3月)该河段各水质参数数据(平均值±标准差)为:水温(13.21±2.24)℃;溶解氧(9.30±0.44)mg/L;pH 7.51±0.16;电导率(60.01±1.15) μS/cm;浊度(1.36±0.11)NTU。
行为观察试验区(图1)采用在自然河道中建坝围网的方法进行构建:选取具有代表性的自然河段(具流水、底质为砾石粗砂、浅滩,比降约5‰),在试验区上下边界建设石坝(坝高高出水面约0.5 m)收拢河道水流以形成急流环境,基于石坝竖立围网(网片孔径为4目;布设范围为自水底至高出水面1.5 m)。为提高试验区水深,在下区尾端石坝水下部分紧贴网片使用PE塑料薄膜控制水流水量。试验区中间布设石坝并预留鱼道,得上下两区:上区约120 m2,水深25~45 cm,为常见的流水浅滩环境;下区水深45~85 cm,为自然河道深水区。
1.2 试验方法
共进行了两次产卵行为观察,分别为预观察(2014年12月-2015年3月)和正式观察(2015月12月-2016年3月)两部分。所有用于观察的试验鱼在培育和产卵观察试验过程中均未注射激素。预观察期间,选取5~7龄的成熟亲本56对放入到试验区内,并采用水下视频监控系统(cy-d3308-16,最大肉眼可辨识视野面积约5 m2)进行行为观察,记录短须裂腹鱼的繁殖期、活动范围、行为类型、产卵方式以及主要活动区域等信息。
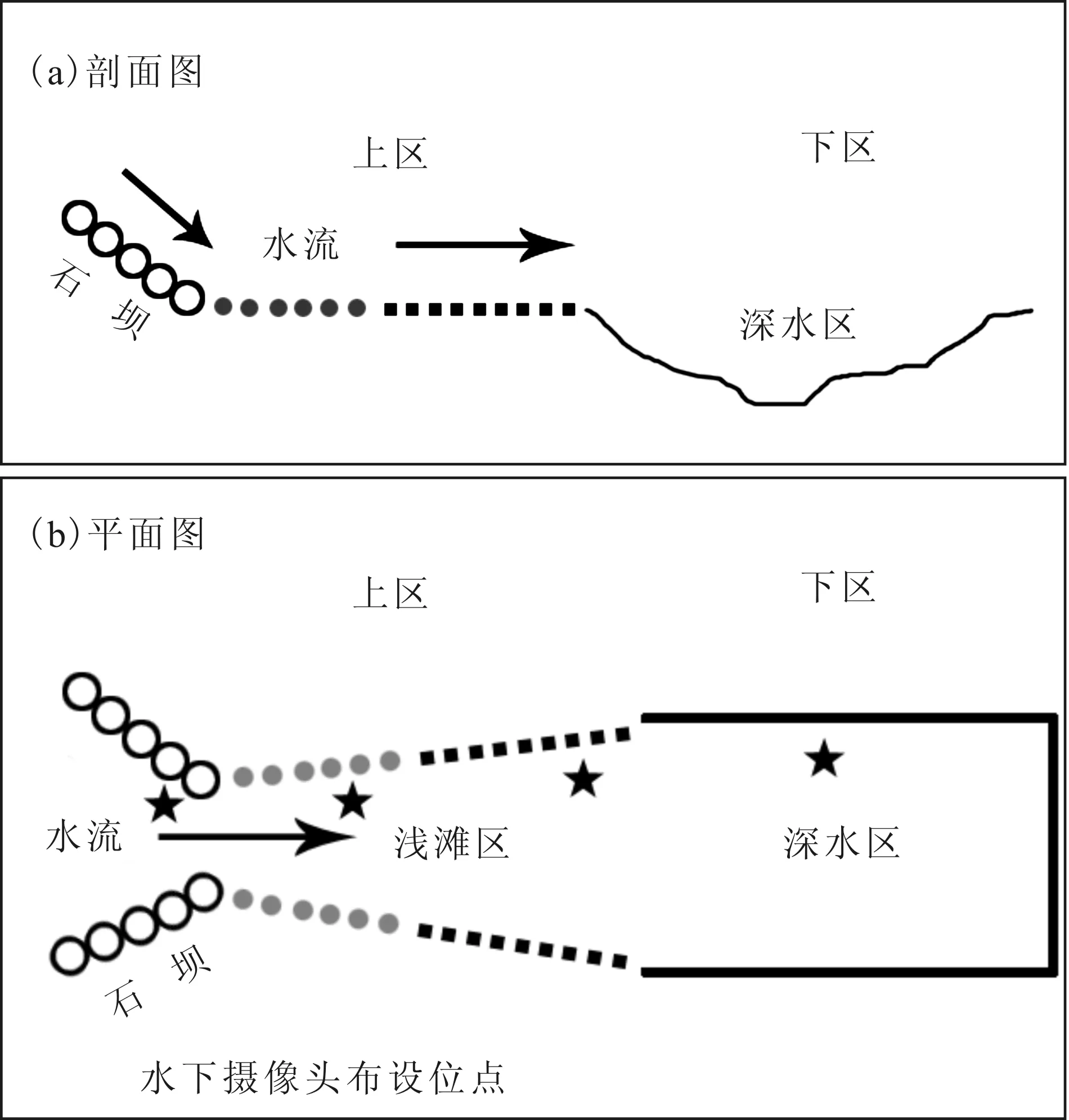
图1 短须裂腹鱼产卵行为观察试验区示意图Fig.1 Diagrammatic map of the experimental site of spawning behavior observation of S.wangchiachii
正式观察期间,选取6~8龄的成熟亲本(雌鱼全长41.2~54.4 cm,体重0.85~1.90 kg;雄鱼全长35.5~52 cm,体重0.55~1.55 kg)35对作为试验鱼投入河道进行观察。根据预观察的结果,在鱼群活动活跃区域布设水下摄像机八台进行连续行为观察和录制。同时使用Sony RX100数码相机在岸边辅助观察记录。所有图像资料均存储于移动硬盘中,以便进一步分析。
采用目标动物取样法和全事件取样法[15]研究短须裂腹鱼产卵行为模式。目标动物取样法:基于预观察,确定核心观测位点及观察的目标行为,采用原位行为摄像的方式对一定时间内某一个体或群体进行连续观察,以获得完整的产卵行为过程。全事件取样法主要观察确定产卵行为发生时间、雌雄鱼产卵行为组成及差异性、各行为特征等。
水下摄像机观察记录时间为07∶00-19∶00,每天均进行记录。水下摄像机主要设置在雄鱼领域、潭区等核心位置。预观察发现,把水下摄像机布设在鱼群固定活动上游位置,拍摄轴线与水流保持一定角度可以获得最佳的拍摄资料;对于通道式的位置,以垂直水流方向水平放置摄像机,可以对巡游或经过的个体进行有效地纪录。为获得最大的水下观察视野,采用高机位俯拍,略低于水平面;同时,水下摄像机布设时尽量避开阳光直射,以保证成像效果。另外,对于发生在水下摄像机拍摄区域以外或活动范围较大的行为,采用岸边观察的方式进行辅助研究。
另外,为了解短须裂腹鱼的繁殖生态特征,在2015-2016年繁殖期整个繁殖期间共解剖测量了20对同来源的暂养于流水池塘中的其他非观察用材料亲鱼,记录了全长、体长、体重、空壳重、性腺重、性腺发育期和绝对繁殖力等指标,并计算了性腺成熟系数[16]。
2 结果
2.1 繁殖生态特性
观察到短须裂腹鱼的繁殖时间为每年12月至次年3月。12月中下旬开始在河边的缓水浅滩、洄水湾即可观察到少量短须裂腹鱼幼苗,同期解剖亲鱼发现:雌鱼性成熟个体达到80%以上,雄鱼100%性成熟;次年3月中旬,亲本性腺已经部分排空,少量未排空的性腺出现退化迹象。
短须裂腹鱼为雌雄异体异型,繁殖期具有明显的第二性征。雄鱼性成熟个体吻部具有大量明显的珠星,通体较为粗糙;雌鱼腹部膨大,具有明显的轮廓,生殖孔突出呈现鲜红色,通体较为光滑。观察试验中可以通过第二性征有效地区分性成熟的雌鱼和雄鱼。
雌鱼个体绝对繁殖力范围为5 150~21 180粒;雌鱼GSI为(13.57±1.24)%,雄鱼GSI为(5.54±0.73)%。
2.2 产卵行为
参照尚玉昌[17]对动物行为的分类方法,依行为发生顺序将试验鱼的产卵行为分为:探索行为、占据与争夺领域、配偶选择、交配行为、亲体护卫等行为。雄鱼参与以上全部行为,雌鱼仅参与配偶选择和交配行为。
2.2.1 探索行为
雄鱼放入河道后立即表现出上行行为。在试验初期雄鱼会对试验场全水域,特别是急流浅滩区域进行巡游。试验最初数天巡游时间可占取样观察时间的60%以上,并且表现出对水流明显的正趋向性。同时在试验场入水处的石坝下存在大量顶水跳跃行为。随着观察试验进行,雄鱼顶水跳跃行为逐渐减少。雄鱼在急流浅滩区域的探索行为会进一步演变为占据行为。
雌鱼放入河道后倾向于顺水而下。尽管雌鱼也会对环境进行探索,但探索区域以深潭、缓水为主。在后期观察中发现,雌鱼探索的区域往往成为产卵期间的休息区域。
2.2.2 领域选择与占据
雄鱼在探索环境的过程中,会挑选适宜产卵的领域并占据。一般选择的领域多为急流浅滩区域,如试验区上游水坝下、试验区上下区连接口处和石块交错下水区。试验观察发现这些区域往往适合产卵,是潜在的产卵点。
雄鱼在占据领域的过程中一般会呈现休息、巡游和顶水三种行为。休息时,全鱼栖于水底,胸鳍张开保持平衡;巡游时,在领域范围内四处游动,驱赶来犯雄鱼及敌害;顶水是指雄鱼在领域内的水层上方顶着水流游动。雄鱼占据的领域一般为1.5~5.0 m2,地形复杂的领域比平缓开阔的的领域小些。为了吸引雌鱼,雄鱼偶尔还会用尾柄打扫领域内卵石上的淤泥,使其保持光洁。
试验场内适合产卵的区域不能满足所有雄鱼占据的需要,因此有些距离较近且中间没有障碍的区域可能为一尾雄鱼占据,也可能是两尾雄鱼在不断争斗的过程中达到一个动态平衡,即各自占据一块,相互之间互不侵犯,当有鱼进入共同区域时随机进行驱赶。当数尾至十数尾雄鱼聚集在同一领域时,往往出现激烈打斗或和平共处的行为表现。对于没有占据领域的雄鱼,多分布在休息区和被其他雄鱼占据的领域周边,伺机争夺领域及跟随雌鱼伺机交配。
2.2.3 领域的争夺与防卫
随着有更多的雄鱼适应环境,雄鱼对有限领域的争斗也越来越明显。争斗行为的方式和强度随着越来越多雄鱼开始性成熟乃至发情,变得越来越激烈,由“威慑”、“驱赶”逐渐变为“打斗”(表1和图2)。
雄鱼防卫的区域一般由内到外可以分为三层(图3):产卵点、巡游区和影响范围,其中产卵点和巡游区两层为领域,面积约为3 m2。但雄鱼防卫领域时,并不局限于领域内,往往提前离开领域对来犯的雄鱼进行驱赶,雄鱼在防卫领域时的活动范围往往越平坦的区域防卫面积越大,一般可达领域面积的3~10倍,这一区域为雄鱼防卫领域时的影响范围。一般情况下雄鱼防卫的核心领域互不重叠,但是影响范围存在一定的重叠。但当数尾雄鱼领域相距较近时,往往会存在一个共同的影响范围,形成一个合作的小群体,在该范围内共同对抗外来雄鱼及敌害物种。
![]()
表1 短须裂腹鱼雄鱼领域防卫模式Tab.1 Male S.wangchiachii territory defensing pattern
![]()
图2 短须裂腹鱼雄鱼的领域防御行为Fig.2 Male S.wangchiachii are defending the territory 左上,一尾雄鱼正在驱赶一尾进入其领域的雄鱼;右上, 一尾守卫领域的雄鱼受到另一尾雄鱼的攻击;左下右下, 一尾雄鱼对一尾多次进入其领域的雄鱼进行连续攻击。
雄鱼守卫领域的时间一般为数小时至数日不等,当领域中存在产卵点,且有雌鱼前来交配时,守卫时间会延长至数周。在产卵高峰期,观察到一尾占据约3.5 m2领域的雄鱼在1 h内防卫“来犯之鱼”143次,其中激烈打斗35次、受到攻击13次、与雌鱼交配3次、整理卵粒14次,其间的巡游时间和休息时间共约14 min。
2.2.4 配偶选择
观察显示,短须裂腹鱼雄鱼比雌鱼发情时间更长。当雌鱼发情时,离开休息区,顶着水流进入急流浅滩寻找合适的雄鱼与产卵点。当雌鱼接近雄鱼的领域时,雌雄双方便会评估彼此的情况决定是否交配。
![]()
图3 短须裂腹鱼领域行为示意图Fig.3 Schematic diagram of S.wangchiachii territory behavior
对于进入自己领域的雌鱼,雄鱼会迎上前主动“展示”自己,围绕雌鱼游动并伴有潜尾行为(雄鱼在围绕雌鱼游动时,多发生下潜趋近并接触雌鱼泄殖孔的行为)和追逐行为。一般雌雄选择耗费的时间较短,数十秒至数分钟之内即可完成。此时没有占据领域的雄鱼也会伺机前来竞争雌鱼,但多被占据雄鱼驱赶离开。当雌鱼确定雄鱼满足配偶要求时,会引领雄鱼到达附近适合产卵的位置,交配产卵(图4)。
当雌鱼认为雄鱼不合适或者其占据的领域不适合产卵时,则会进一步寻找并评估其他雄鱼及其领域。此时被淘汰的雄鱼往往会继续跟随雌鱼,由占据领域的优势者变成其他领域的偷袭者。雄鱼在这一过程中也会不断调整其占据领域的范围和类型,以获得更多雌鱼的选择。
2.2.5 交配产卵
观察显示短须裂腹鱼为分批产卵型。产卵行为均分在繁殖期中,且单日可以多次产卵(曾观测到同一尾雌鱼一天产卵8次,间隔(34.14±16.85)min)。观察到的产卵点多为水深25~55 cm、水流湍急的沙石滩。当雄鱼获得雌鱼的认可后,雌鱼会巡视雄鱼的领域,并在确定合适的产卵位点后引领雄鱼至该位点进行产卵。
![]()
图4 一尾短须裂腹鱼雌鱼正在引领一尾雄鱼进入产卵点Fig.4 A female S.wangchiachii is leading a male fish into the spawn point
雌鱼躯干上扬尾部弯曲上翘,臀部贴地并剧烈地摆动,形成一沙窝;雄鱼紧贴雌鱼,躯干微微上仰,尾部剧烈抖动(图5)。该行为确定为交配行为。在雌鱼排卵行为发生时,附近伺机的雄鱼会采用“偷袭”的方式前来参与交配:贴近交配的雌鱼和雄鱼两边,趋近雌鱼泄殖孔,同交配中的雄鱼一样,躯干微微上仰,尾部剧烈抖动。观察发现其中一尾鱼的交配持续时间为7~28 s。
![]()
图5 一对短须裂腹鱼亲鱼的两次产卵行为Fig.5 Two times of spawning behavior of the sameS.wangchiachii parents in the same territory
由于占据此领域的雄鱼正在与雌鱼交配,无暇驱赶偷袭者,从而形成多个雄鱼与一尾雌鱼交配的现象。实际观察中,发现交配的雌雄性比多为1∶2和1∶3,在雄鱼发情期高峰时会有更多的雄鱼参加。
雌鱼有时尾柄抖动时间少于7 s即停止,然后重新找一处合适的位置再一次产卵,则之前的抖动行为可认为是伪产行为。伪产位点不存在或者极少存在卵粒,而正常的产卵点通过实地考察可以找到200~500粒卵。
2.2.6 亲体护卫
完成交配后的雌鱼随水流而下回到休息区,雄鱼则开始检查产卵情况,对没被砂石埋藏的卵粒进行埋藏。埋卵时,雄鱼首先用口含取就近的沙粒,再吐在位于凹处的暴露卵粒上(图6),对位于平整位置的暴露卵粒则通过身体摆动引导水流把卵粒冲到下游石砾缝隙中。此过程参与交配的雄鱼都有参与。埋卵完成后除占据领域的雄鱼外,其他雄鱼渐渐离开。
占据此领域的雄鱼会一直在产卵点附近进行护卫。对产卵点的护卫行为和交配前的领域防卫行为基本类似。区别在于,雄鱼对产卵点区域更为敏感,往往对靠近产卵点的个体与群体作为优先驱赶的对象。同时发现雄鱼在休息时往往避开产卵点,这可能是雄鱼为了防止产卵点的暴露。对于产卵点中重新裸露出来的卵粒,雄鱼会及时进行埋藏,并且清理卵粒附近的污物。
![]()
图6 一尾雄鱼正在含取沙粒掩盖卵粒Fig.6 A male S.wangchiachii is covering up the eggs with sand by its mouth
2.3 其他行为
2.3.1 休息行为
雌鱼在发情以外的时间内几乎都处于休息状态,栖息于静水深潭和缓水区。休息期间,雌鱼一般趴在水底砂石上,或藏身于水底石块缝隙中,或迎着缓水流动方向顶水游动。雄鱼因长时间处于领域争夺与防卫行为中,只有在环境不适如温度过高、光照强度过高以及存在人为干扰的情况下,才会回到深水区躲藏或休息。
2.3.2 摄食行为
试验期间,多次观察到试验鱼的摄食行为,主要分为捕食(发现水面有潜在食物时,通过身体上仰、冲刺和衔食等行为,实现对水面漂浮的藻类、小型昆虫及投喂饲料的摄取)和啃食(摄食水底的有机碎屑及着生藻类时,采用刮、咬、撕扯等动作完成摄食)两种行为。
2.4 产卵行为模式
根据各个行为的发生条件和发生顺序,建立了短须裂腹鱼产卵行为模式。短须裂腹鱼繁殖期间行为的多样性以及复杂性主要体现在雄鱼的行为上。繁殖期到来时,雄鱼首先通过探索行为,寻找适合产卵的环境。适宜产卵的空间环境是有限的,因此雄鱼会为了获得优质产卵环境而进行不断地探索与争夺。没有获得领域的雄鱼,要么潜伏在有雄鱼占据的领域周边伺机争夺领域,要么在休息区的周边等待着跟随顶水上行产卵的雌鱼进行“偷袭”交配。雌鱼发情后,开始顶水上行至急流浅滩区域,评估和选择恰当的雄鱼进行交配。雌鱼主要通过评估雄鱼占据领域的质量、身体状况等来确定与之交配的对象。确认交配时,雌鱼会引导雄鱼至合适的产卵点进行交配产卵,此时常有伺机而动的雄鱼加入到交配活动中。而占据雄鱼为了顺利完成交配,一般无暇驱赶偷袭者。产卵完成后,雌鱼即行离开,雄鱼则立即检查和填埋暴露在外的卵粒,并护卫产卵点。参与偷袭的雄鱼,也会参与暴露在外卵粒的填埋工作,但完成后即离开,不参与产卵点的护卫。
3 讨论
鱼类产卵行为是遗传调控和环境选择共同作用的结果,生活在不同环境的鱼类有着多种多样的产卵行为模式(表2)。短须裂腹鱼生活在长江上游干支流中,其典型的产卵行为特征如雄鱼不筑巢、雄鱼排沙埋卵、雌鱼分批产卵,是有助于其适应环境的。
有关裂腹鱼类产卵繁殖习性的报道极少。陈永祥等[18]曾对四川裂腹鱼的繁殖习性进行调查研究,其通过走访渔民和采集亲鱼样本等调查后认为,四川裂腹鱼存在集群行为,产卵前有追逐行为,这与本研究揭示的短须裂腹鱼产卵行为特征大体类似。但陈永祥等[18]根据四川裂腹鱼雄鱼腹部有伤痕、产卵点存在沙窝等特征,认为四川裂腹鱼雄鱼存在筑窝习性,这与本研究发现的短须裂腹鱼雄鱼不筑巢是明显不同的。在本研究中,短须裂腹鱼雄鱼在打扫领域时会用尾柄摩擦卵石,但其覆盖面积较广,不会集中一点挖成一个沙窝,而雌鱼在产卵时会用尾柄贴地抖动形成沙窝。由此可见,陈永祥等有关四川裂腹鱼雄鱼存在筑窝习性的证据,实际上有可能是限于技术条件,误把雌鱼产卵形成的沙窝认为是雄鱼造成的,当然这还需要直接的观察试验来验证。
很多在流水浅滩环境中产卵的鱼类都会有筑巢的行为,如七鳃鳗[19]、三刺鱼[17]和鲑亚科鱼类[8]。筑巢有助于展示雄鱼的繁殖能力,提升雄鱼对雌鱼的吸引力[20]。而短须裂腹鱼在产卵过程中不筑巢的原因可能有三点:一是短须裂腹鱼雌鱼产卵点并不固定,雄鱼往往是在雌鱼的带领下进入特定的产卵位点;二是短须裂腹鱼生存环境中还存在大量的天敌,如双斑锦蛇,把卵粒集中到一处巢穴中易被捕食者识别,增加了后代被捕食的风险;三是短须裂腹鱼受精卵的发育时间较短,一般为119~192 h[21],筑巢的回报可能较低,而选择将更多的能量投入到亲体护卫行为中,可能会有更高的回报。
排沙埋卵是比较少见的鱼类产卵行为特征。目前,还未见有关鱼类存在亲鱼埋卵行为的研究报道。同时,由于缺乏其它鱼类在野生条件下直接观察产卵行为的报道,当前还无法确认排沙埋卵是否是短须裂腹鱼特有的产卵行为特征。短须裂腹鱼埋卵行为的作用可能有以下三点:一是防止捕食者对卵粒的捕食或破坏;二是为受精卵提供良好的孵化条件,如隔绝强紫外线照射、防止淤泥粘染;三是防止水流将受精卵冲到下游不适于孵化的环境中。
短须裂腹鱼属分批产卵鱼类。本研究中,短须裂腹鱼雌鱼绝对繁殖力为5 150~21 180粒。但在交配行为发生后,产卵点仅能收集到200~500粒。同时,行为观察过程中发现,同一尾雌鱼一天之内也会发生数次交配行为,雌鱼的每一次发情都会重新选择雄鱼与产卵点。由此可以推断,短须裂腹鱼属分批产卵鱼类,且每次产卵都会重新选择交配对象和产卵点。
本研究是在近乎自然的环境中对人工驯养后的短须裂腹鱼亲鱼进行初步的产卵行为观察,其观察结果可在一定程度上揭示裂腹鱼类的自然产卵行为特性,但是要想进一步获得自然环境中野生裂腹鱼类的产卵行为,还是需要进一步的技术改进和深入的观察研究。
![]()
表2 几种常见鱼类产卵行为模式的比较Tab.2 The comparison of spawning behavior patterns among several different fish species
注:“—”表示资料不详。
[1]丁瑞华.四川鱼类志[M].成都:四川科学技术出版社,1994:365-367.
[2]乐佩琦.中国动物志:硬骨鱼纲鲤形目(下卷)[M].北京:科学出版社,2000:291-292.
[3]高少波,唐会元,乔 晔,等.金沙江下游干流鱼类资源现状研究[J].水生态学杂志,2013, 34(1):44-49.
[4] 刘跃天,冷 云,徐伟毅,等.短须裂腹鱼人工繁殖初探[J].水利渔业,2007,27(5):31-32.
[5] 李光华,冷 云,吴敬东,等.短须裂腹鱼规模化人工繁育技术研究[J].现代农业科技,2014,10:259-261,270.
[6] 刘汉生,易祖盛,林小涛.唐鱼的繁殖行为和胚胎发育研究[J].水生态学杂志,2008,1(2):22-27.
[7] 谢增兰,胡锦矗,郭延蜀,等.叉尾斗鱼繁殖行为的观察[J]. 动物学杂志,2006,41(5):7-12.
[8]EsteveM.ObservationsofspawningbehaviourinSalmoninae:Salmo,OncorhynchusandSalvelinus [J].RevFishBiolFisher,2005,15(1-2):1-21.
[9]TheriaultV,MoyerGR,JacksonLS,etal.Reducedreproductivesuccessofhatcherycohosalmoninthewild:insightsintomostlikelymechanisms[J].MolEcol,2011,20(9):1860-1869.
[10]Venditti,D A, James C A.Reproductive behavior and success of captive-reared chinook salmon spawning under natural conditions[J].N Am J Fish Manage,2013,33(1):97-107.
[11]Teresa F B, Freitas E G.Reproductive behavior and parental roles of the cichlid fishLaetacaraaraguaiae[J].Neotrop Ichthyol,2011,9(2):355-362.
[12] Rezucha R,Smith C, Reichard M.Personality traits,reproductive behaviour and alternative mating tactics in male European bitterling,Rhodeusamarus[J].Behaviour,2012,149(5):531-553.
[13]Nanami A,Sato T,Ohta I,et al.Preliminary observations of spawning behavior of white-streaked grouper (Epinephelusongus) in an Okinawan coral reef[J].Ichthyol Res, 2013,60(4):380-385.
[14]蒋志刚.动物行为原理与物种保护方法[M].北京:科学出版社,2004,1-16.
[15] 乔征磊,张洪海,马建章,等.半散养东北虎繁殖期PAE编码行为谱的构建[J].生态学杂志,2015,34(3):736-743.
[16]殷名称.鱼类生态学[M].北京:中国农业出版社,1995:111-112.
[17]尚玉昌.行为生态学[M].北京:北京大学出版社,1998:105-110.
[18] 陈永祥,罗泉笙.四川裂腹鱼繁殖生态生物学研究Ⅴ、繁殖群体和繁殖习性[J].毕节师专学报,1997,(1):1-5.
[19]Li W,Scott A P,Siefkes M J.et al.Bile acid secreted by male sea lamprey that acts as a sex pheromone[J].Science,2002,138(296):138-141.
[20] Lehtonen T K,Lindstrom K.Females decide whether size matters: plastic mate preferences tuned to the intensity of male-male competition[J].Behav Ecol,2009,20(1):195-199.
[21]刘 阳,朱挺兵,吴兴兵,等.短须裂腹鱼胚胎及早期仔鱼发育观察[J].水产科学,2015,34(11):683-689.
[22]廖彩萍.高体鳑鲏繁殖行为生态学研究[D].北京:中国科学院研究生院,2012:23-28.
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼第一线"微信公众帐号和微信视频号"养鱼第一线刘文俊视频号"以及头条号"水花鱼@渔人刘文俊"!
