


通讯:13983250545
微信:ycsh638
QQ:469764481邮箱:ycsh6318@163.com
论文:齐口裂腹鱼感染嗜水气单胞菌后血液生化指标与组织病理学研究

齐口裂腹鱼感染嗜水气单胞菌后血液生化指标与组织病理学研究
罗 辉1,2,3,杨月静2,向梦斌2,叶 华2,郑曙明2,王晓清1,3
(1.湖南农业大学动物科学技术学院,湖南长沙 410128;2.西南大学鱼类繁育与健康养殖研究中心,重庆 402460;3.水产高效健康生产湖南省协同创新中心,湖南常德 415000)
为了研究嗜水气单胞菌(Aeromonashydrophila)对齐口裂腹鱼(Schizothoraxprenanti)血液生理生化指标的影响,将180尾齐口裂腹鱼平均分成6组(4、24和48 h对照组;4、24和48 h感染组)。对照组腹腔注射0.2 mL 0.65%的无菌生理盐水,感染组腹腔注射0.2 mL 2.5×107CFU/mL的嗜水气单胞菌液。分别测定4、24和48 h对照组和感染组齐口裂腹鱼各项血液生理生化指标,并对肾脏、肝脏和肌肉组织进行病理学观察。结果显示:谷丙转氨酶、白蛋白、总胆固醇、葡萄糖、乳酸脱氢酶、钾、钠、氯、钙和磷含量均有显著变化。谷草转氨酶、碱性磷酸酶、总蛋白、球蛋白、甘油三酯则没有受到显著影响。由此可见,嗜水气单胞菌感染后对齐口裂腹鱼血液指标产生了显著的影响。对病鱼的病理学观察发现病鱼体表多处出现充血出血,溃疡和腹水,尤其是肝脏、肾脏和肌肉组织均出现不同程度的损伤,变性和局部坏死。
齐口裂腹鱼(Schizothoraxprenanti);嗜水气单胞菌(Aeromonashydrophila);血液生理生化指标;组织病理
齐口裂腹鱼(Schizothoraxprenanti(Tchang))是长江上游著名的土著经济鱼类,在分类上属于鲤形目鲤科裂腹鱼亚科裂腹鱼属。齐口裂腹鱼为底栖性冷水鱼类,其肉质细嫩,肉味鲜美,极具商业价值[1]。其人工养殖和繁殖的相关研究在2001年取得初步成功后[2-4],逐渐在四川、重庆、云南等地形成了规模养殖[5]。伴随着养殖规模和密度的迅速扩大,齐口裂腹鱼的养殖业也饱受细菌和寄生虫病的困扰,其中尤以嗜水气单胞菌等为代表的细菌性疾病以其高致病性,高死亡率和难治愈性危害极大[6-10]。嗜水气单胞菌曾在四川雅安和攀枝花等地恶性传染,导致养殖鱼30%~80%的感染,病鱼体表严重损伤,肌肉溃烂,从而失去商品价值[8]。
嗜水气单胞菌(Aeromonashydrophila)是鱼类常见的重要病原菌,可引起组织局部损伤、坏死、水肿、突眼、腹部膨胀、腹水、贫血以及内脏器官损伤等多种症状,给水产养殖业带来巨大的经济损失[11,12]。研究表明嗜水气单胞菌感染会引起水产动物血液生理生化指标发生变化,尤其对免疫功能和肝功能相关生化指标产生显著影响[5,13-14]。然而目前关于嗜水气单胞菌感染后对齐口裂腹鱼血液生化指标和组织病理损伤影响的研究较少[5,15]。病原菌侵袭水生动物后,在进入体内的最初0~24 h通常为其粘附宿主组织并缓慢扩增期。在成功定植后,24~48 h即可达到指数增长期,48 h以后细菌的增殖就进入平台期[16]。因此,如何在感染早期确诊就显得尤为重要,其研究成果对鱼类的人工养殖和疾病防治更具指导意义。但嗜水气单胞菌感染后在细菌缓慢扩增期(0~24 h)和指数增长期(24~48 h)对齐口裂腹鱼血液生理生化指标和组织病理学的影响仍未见报道。本实验通过对嗜水气单胞菌感染后(0~48 h)齐口裂腹鱼的外观症状及组织病理学观察,结合4、24和48 h三个时点分别设置感染组和对照组的齐口裂腹鱼血液生理生化指标的测定,初步探讨了嗜水气单胞菌对齐口裂腹鱼组织损伤和血液生理生化指标的影响。本研究可为齐口裂腹鱼后续疾病防治、免疫机制和种质资源保护等相关研究提供理论基础和数据积累。
1 材料与方法
1.1 实验材料
本实验于2014年10—11月在西南大学鱼类繁育与健康养殖研究中心进行。实验材料为人工养殖的齐口裂腹鱼,体重(105±36)g,体长(19±2)cm。试验鱼经过2周的暂养,停食1 d后选择体质健壮,反应灵敏的个体进行攻毒实验,攻毒后停止饲喂。
1.2 实验方法
1.2.1 细菌培养
本研究所用嗜水气单胞菌为西南大学鱼类繁育与健康养殖研究中心鉴定保存的菌株,分离自患典型细菌性败血症的南方鲶内脏。将嗜水气单胞菌菌株在LB培养基于28 ℃条件下培养24 h,然后挑取单菌落于LB培养基,28 ℃,200 r/min培养过夜,取150 μL菌液涂布到无氨苄的LB固体培养基上,在28 ℃下培养6 h,用无菌生理盐水收集菌液,用麦氏比浊法测定细菌数量,再用无菌生理盐水稀释至2.5×107CFU/mL ,置于4 ℃保存备用。
1.2.2 攻毒试验
选择180尾健康无伤的暂养齐口裂腹鱼,分成2组,每组3个重复(3个重复分别对应取样时间4、24和48 h),每重复30尾鱼。第一组为对照组,腹腔注射0.2 mL 0.65%的无菌生理盐水;第二组为攻毒组,分别腹腔注射0.2 mL 2.5×107CFU/mL的嗜水气单胞菌,每组注射结束后记录时间。保持水温在20 ℃±1 ℃,增氧机持续增氧。
1.2.3 样品的采集与测定
在注射无菌生理盐水或嗜水气单胞菌后4、24和48 h分别在对应重复组中随机取5尾鱼,用300 mg/L的MS222麻醉,用2 mL一次性无菌注射器尾静脉取血,采血后测量体长、体重,随后迅速解剖鱼体,取肌肉、肝脏和肾脏用于组织病理学观察。血液迅速放入离心管内,2 000 r/min离心10 min,用移液枪将上层澄清血液转入PT离心管中备用。每尾鱼的血液生化指标及离子成分的测定采用北京九强生物技术有限公司生产的试剂盒,在自动生化分析仪(Beckman Coulter,USA)测定(其中酶活单位的定义为37 ℃条件下,每分钟转化1 μmol/L底物的酶量为1个酶活单位,即为U/L)。分别取对照组和有典型症状的齐口裂腹鱼的肝脏、肾脏和肌肉组织,用Bouin氏液固定24小时,在经过脱水和透明处理后用石蜡包埋。通过KD-202轮转切片机切片后HE染色,用中性树胶封片,在正置DPT2型OLMPUS显微镜拍摄系统下观察并进行显微拍照。
1.3 数据处理
实验所得数据用平均值±标准差(Mean±SD)表示。对照组数据取注射无菌生理盐水三个时点(4、24和48 h)的平均值,注射嗜水气单胞菌的各时点数值经过标准化处理,公式为Y=个体某一指标测定值 ×(对照组对应指标三个时点的平均值/单一时点某指标平均值)。采用SPSS 20.0统计软件进行数据统计分析。采用单因素方差分析和Duncan法多重比较各组的差异显著性。
2 结果与分析
2.1 攻毒后外观症状
腹腔注射嗜水气单胞菌后4 h观察齐口裂腹鱼与对照组无明显差异,游动正常,外观无明显症状。24 h开始出现明显发病症状,主要表现为鱼体发黑,体表粘液分泌增加,部分鱼游动急促,呼吸加快,继而呆滞不动,体表和鳍条基部可见充血发红,并出现1尾死鱼。48 h死鱼数量增至15尾,鱼体表出现溃疡,充血更加明显,病鱼出现运动失调,无法保持平衡。临检病鱼可发现体表多处充血,体表粘液增多,眼球微微外凸,腹部膨大,腹壁变薄,上下颌及颐部严重充血(图1),鳍条及鳍条基部充血,肛门红肿外凸。解剖后腹腔内有大量黄色或带血积水,肝、肾、脾等内脏充血甚至肿胀,严重者肠出现糜烂(图2)。

图1 发病鱼体表充血、溃疡(←)Fig.1 Hyperemia and ulceration on the body surface of the diseased fish(←)

图2 发病鱼内脏充血、肿胀(←)Fig.2 The diseased fish showed enlarged and reddened viscera(←)
2.2 嗜水气单胞菌感染对齐口裂腹鱼血液生化指标的影响
嗜水气单胞菌感染后齐口裂腹鱼血液生化指标的测定结果见表1。感染组48 h齐口裂腹鱼血液中谷丙转氨酶浓度显著高于对照组,同时也显著高于4 h和24 h组。血液中谷丙转氨酶浓度随着时间的延长而升高,尽管24 h与4 h组之间差异不显著,但升高的趋势明显。攻毒后4 h,齐口裂腹鱼血清中白蛋白含量显著低于对照组,也低于24 h和48 h组,但白蛋白随着攻毒后时间的延长逐渐升高至与对照组差异不显著的水平。攻毒的三个时间点血清中总胆固醇含量与对照组相比,差异均不显著,但4 h与48 h两组间则差异显著,从其变化规律可以看出,随着攻毒后时间的延长,血清中胆固醇含量逐渐回升,并在48 h时显著高于4 h组,其余各组间则差异不显著。攻毒后48 h血糖含量显著低于对照组,其余两组与对照组差异不显著,且攻毒后血糖含量随着时间的延长,逐渐降低,但相互之间没有显著差异。攻毒后各组乳酸脱氢酶含量与对照组差异均不显著,但48 h组显著高于24 h组。
攻毒后血清中谷草转氨酶、碱性磷酸酶、总蛋白、球蛋白和甘油三酯的含量与对照组均无差异显著性,且攻毒组各时点之间差异也不显著。但从各指标的数据可以看出,碱性磷酸酶、总蛋白和球蛋白在攻毒后存在一定的规律性变化,即在4 h时血清中含量最低,并随着时间的延长,有逐渐升高的趋势。而甘油三脂的变化则是随着时间的延长有逐渐降低的趋势。
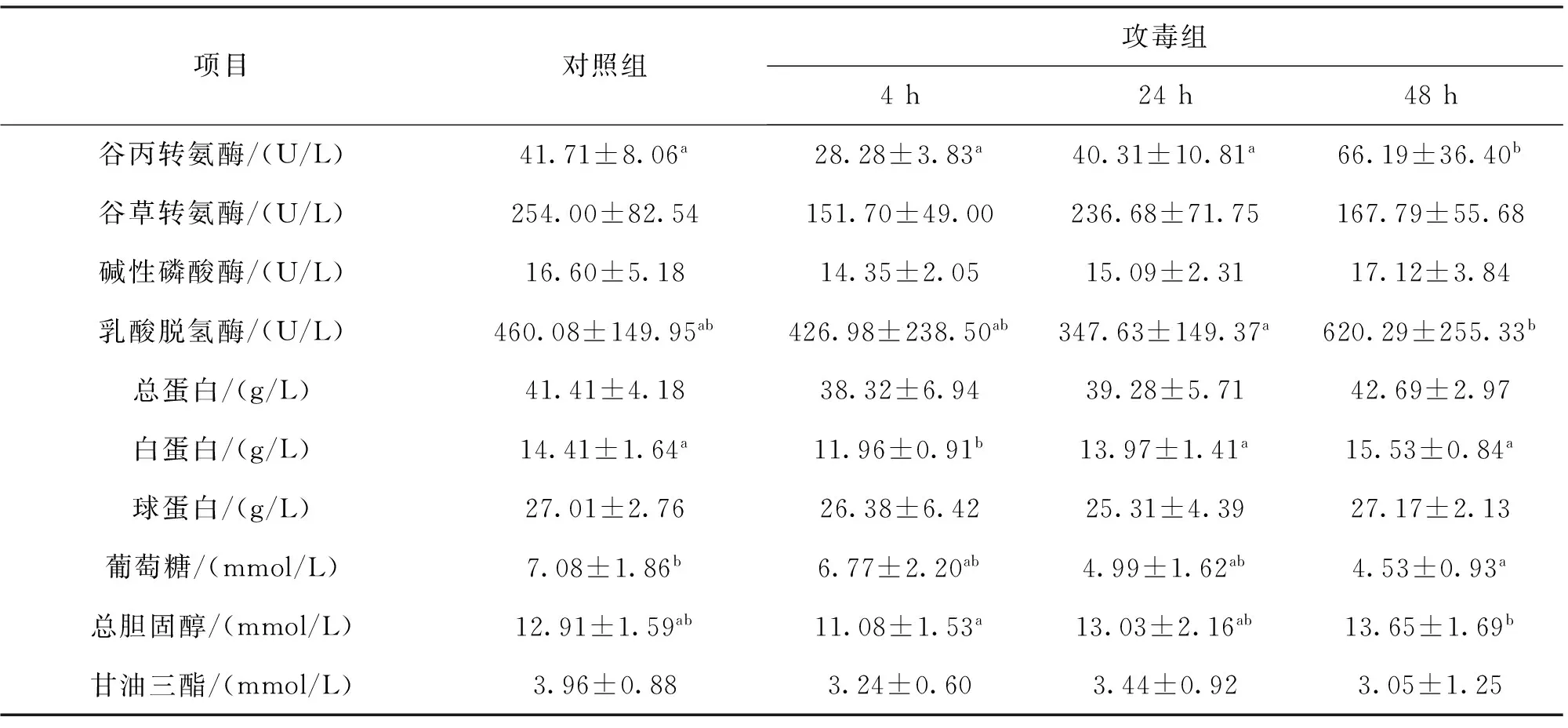
表1 嗜水气单胞菌对齐口裂腹鱼血液生化指标的影响Tab.1 Effect of A.hydrophila on blood biochemical indices of S.prenanti
注:同一行数据不同上标字母表示差异显著(P<0.05)。表2同。
2.3 嗜水气单胞菌感染对齐口裂腹鱼血液离子水平的影响
嗜水气单胞菌感染后齐口裂腹鱼血液离子水平的测定结果见表2。攻毒后血液中的离子含量均发生了显著变化。其中钾、钠和氯离子有相似的变化规律,攻毒后24 h钾离子浓度显著高于对照组,其余各组与对照组差异不显著。而钠离子则是在攻毒后4 h显著升高,24 h和48 h血中钠离子水平有所回落,与对照组差异不显著。血清中氯离子浓度也在攻毒后4 h显著高于对照组,但随着时间的延长其含量又逐渐降低至对照组相近水平,最终在24 h和48 h时与对照组差异不显著。血清中钙离子和磷离子的变化趋势类似,即在攻毒后4 h血清中含量显著低于对照组,但随着时间的延长又逐渐升高。其中钙离子水平除在4 h时显著低于对照组外,24 h和48 h含量升高,与对照组差异不显著。磷离子水平则是4 h时显著低于对照组,随着时间的延长逐渐升高,并在48 h时显著高于对照组。
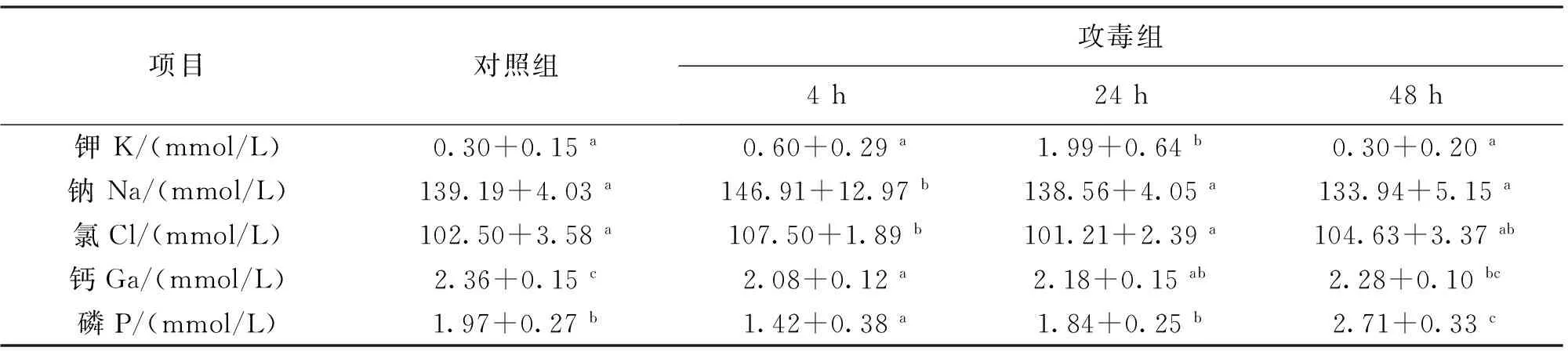
表2 嗜水气单胞菌对齐口裂腹鱼血液离子水平的影响Tab.2 Effect of A.hydrophila on concentration of ion in blood of S.prenanti
2.4 组织病理变化
齐口裂腹鱼感染嗜水气单胞菌的病理学变化主要表现为机体多组织器官充血出血,肿胀,细胞坏死、变性,并伴有炎症。对照组肌肉组织中肌细胞排列紧密,条纹清晰,细胞核位于细胞边缘。病鱼肌细胞着色不均,部分区域肌浆溶解,肌原纤维断裂、弯曲不齐,肌纤维间隙增大(图3)。对照组肾脏肾小管结构清晰,肾间质毛细血管内充满红细胞,各细胞结构正常。病鱼肾脏的肾小管上皮细胞排列松散,管腔结构不清晰,肾间质出血,伴有嗜中性白细胞浸润(图4)。对照组肝脏肝细胞界限清晰,细胞核圆形深染,位于细胞一侧,胞浆不着色空泡化。病鱼肝细胞严重空泡变性,细胞核崩解,细胞融合、界限消失,少数肝细胞出现坏死(图5-6)。

图3 病鱼肌细胞着色不均, 部分区域肌浆溶解,肌原纤维断裂(←)Fig.3 The myocytes of disease fish showed uneven coloring, the sarcoplasm dissolved,the myofibril ruptured
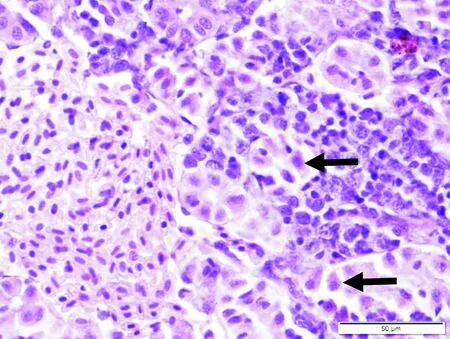
图4 病鱼肾小管上皮细胞 排列松散(←),管腔结构不清,肾间质出血Fig.4 The tubular epithelial cells arranged in loose,the lumen structure is unclear,the renal interstitium suffer a hemorrhage
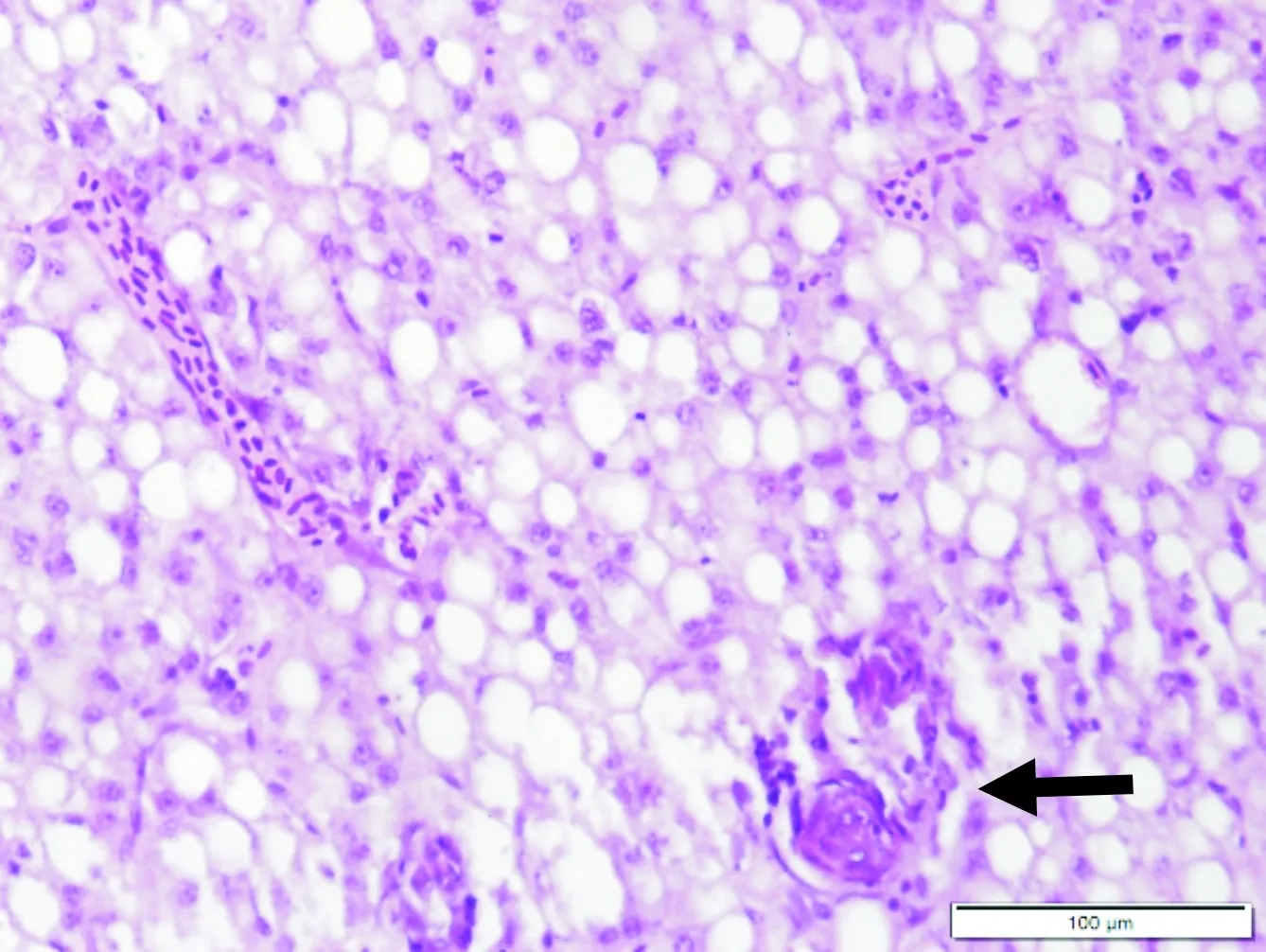
图5 病鱼肝细胞空泡变性,局部区域出现坏死(←)Fig.5 Liver cells of diseased fish were vacuolar degeneration and necrosis in local area(←)
3 讨论
3.1 嗜水气单胞菌对齐口裂腹鱼的致病性
过去通常认为致病性嗜水气单胞菌是条件性致病菌,但近年来的研究表明其更可能是原发性致病菌[7,17],并已发展成为水产养殖的一大杀手,给我国鲤 (Cyprinuscarpio)、鲫 (Carassiusauratus)、黄鳝(Monopterusalbus)、香鱼(Plecoglossusaltivelis)、虹鳟(Oncorhynchusmykiss)、鲈 (Micropterussalmoides)、鳗鲡(Anguillajaponica)、白斑狗鱼(Esoxlucius)、牛蛙(Ranacatesbeiana)、鳖(Trionyxsinensis)等水产动物的养殖带来了严重的损失[17-23]。本实验使用嗜水气单胞菌人工感染齐口裂腹鱼,导致其在24 h全身充血出血,48 h明显出现眼球突出、溃疡等症状,甚至死亡,与文献报道的齐口裂腹鱼自然感染症状相似[7,8],同时与其他淡水鱼类爆发嗜水气单胞菌病的症状也有诸多相似之处[17,22-23]。
3.2 嗜水气单胞菌对齐口裂腹鱼血液生化指标和组织器官的影响
血液中的谷丙转氨酶和谷草转氨酶在正常情况下酶活低且相对恒定,但当肝脏等组织受到损伤或受到环境胁迫时,血液中这两种酶活将升高,二者在评估肝脏和其他器官的状态方面起着重要的作用[24]。本研究发现齐口裂腹鱼感染嗜水气单胞菌48 h时血清中谷丙转氨酶显著高于对照组,同时发现血清中谷丙转氨酶随着感染时间的延长而升高,在48 h时谷丙转氨酶浓度比4 h时升高141%。从肝脏、肾脏和肌肉的病理切片可以看出病鱼肝脏受损,肝细胞严重空泡变性,细胞核崩解,细胞融合、界限消失,少数肝细胞出现坏死。肾脏和肌肉组织也受到破坏,肾间质出血,伴有嗜中性白细胞浸润,病鱼肌肉组织中肌浆溶解,肌原纤维断裂、弯曲不齐,肌纤维间隙增大,提示血液中谷丙转氨酶的升高可能是肝脏等器官受损导致其分泌到血液中所致。这与大黄鱼(Larimichthys crocea)感染哈维氏弧菌(Vibrio harveyi)和欧洲鳗鲡患“狂游病”的研究结果类似[25,26]。
研究表明乳酸脱氢酶的含量与鱼体肌肉组织的健康状况有关[25,27]。徐晓津等[26]研究发现大黄鱼感染哈维氏弧菌后病鱼乳酸脱氢酶含量显著升高,表明大黄鱼肌肉组织受到破坏,这与大黄鱼患病后产生皮肤溃疡的症状相吻合。本研究发现尽管攻毒组在4 h和24 h时乳酸脱氢酶含量与对照组没有显著差异,但48 h攻毒组则显著高于对照组,并显著高于24 h攻毒组。推测攻毒后鱼体肌肉组织受损,产生了病变,这与肌肉病理切片发现病鱼肌浆溶解,肌原纤维断裂、弯曲不齐,肌纤维间隙增大的病理症状相吻合。有趣的是攻毒后24 h组乳酸脱氢酶含量与对照组和攻毒后4 h尽管没有显著差异,但却有较明显的下降趋势,其中的原因有待进一步深入研究。
血中葡萄糖是动物机体生化活动所需能量的来源之一,在正常生理状态下,其含量相对恒定,但会随着机体活动和适应环境因子的改变而变化。因此血糖含量可作为鱼体响应环境应激因子的指示指标[28]。本研究中,齐口裂腹鱼感染嗜水气单胞菌后随着时间的延长,血糖含量显著下降,48 h组血糖含量显著低于对照组。这可能是由于机体在病菌侵入后迅速调动先天性免疫防疫机制以抵抗病原的粘附和增殖,从而消耗大量能量致使血液中血糖含量显著下降。徐晓津等[26]在研究大黄鱼(Larimichthyscrocea)感染哈维氏弧菌(Vibrioharveyi)时,观察到病鱼血液中血糖含量显著下降。余毅等[29]在患疯狂病的白鲢(Hypophthalmichthysmolitrix)血液指标的研究中也发现病鱼血糖含量显著降低,与本研究的结果类似。
血清蛋白在维持血浆正常胶体渗透压,对物质的运输和体内酸碱平衡方面都起到十分重要的作用。因此血清总蛋白、白蛋白和球蛋白含量也可以作为衡量鱼类响应外界应激的重要参考指标[20]。本研究中齐口裂腹鱼感染嗜水气单胞菌后血清总蛋白、球蛋白均没有显著变化,但白蛋白含量在4 h时显著低于对照组。在24 h和48 h时血清中白蛋白含量有所回升,并与对照组差异不显著。总蛋白含量也有攻毒后降低,随后逐渐升高的趋势。分析原因可能是由于机体在病原侵入机体后迅速动用免疫防御机制对病原进行清除,白蛋白作为载体在此过程中被迅速消耗,但随着时间的延长,机体稳态的调节使白蛋白的数量逐步恢复正常。强俊等[30]在研究海豚链球菌(Streptococcusiniae)感染罗非鱼(Oreochromisspp.)时也观察到血清总蛋白、白蛋白和球蛋白从0~48 h期间经历了先降低后升高的变化,与本研究观察到的现象类似。
3.3 嗜水气单胞菌对齐口裂腹鱼肾脏和血液离子水平的影响
鱼类的肾脏不仅是重要的免疫器官,同时也是重要的排泄器官,对调节鱼体渗透压和水盐平衡至关重要[31]。本研究中发现齐口裂腹鱼肾脏明显受损,肾小管上皮细胞排列松散,管腔结构不清,肾间质出血,并伴有嗜中性白细胞浸润。这必然影响鱼体正常的生理代谢和免疫功能。本研究中发现Na+和Cl-含量在感染4 h时显著升高,但随后逐渐降低至与对照组差异不显著。 K+含量也在24 h显著升高,但在48 h也降低到与对照组差异不显著的水平。分析可能是细菌攻击宿主肾脏等组织,导致细胞骨架重排,渗透性异常,从而使血中Na+、Cl-、K+浓度升高。同时细胞骨架重排等效应是短暂且可逆的,重排的骨架蛋白在所附着的病菌下部短暂积聚,待病菌被细胞摄入后,骨架中的某些成分迅速解离,细胞骨架又恢复到稳定平衡状态[32],从而使血清中离子浓度趋于正常。徐晓津等[26]在大黄鱼感染哈维氏弧菌的研究中也发现血浆中K+和Cl-的浓度在感染后显著升高,与本实验结果一致。
研究表明当肾脏受损时,机体磷代谢就会出现异常。在人类慢性肾衰竭时,机体磷代谢发生紊乱,即出现血磷升高[33]。本研究中齐口裂腹鱼感染后4 h血液中磷含量显著低于对照组,而在48 h时显著高于对照组。这可能是病原入侵机体后,磷首先作为ATP、核酸、磷脂、细胞膜和多种辅酶的重要组成成分,参与能量转化等多种生理生化反应而被消耗,但随着时间的延长,病原对鱼体肾脏的损害加剧,导致肾脏功能紊乱,磷排泄异常,致使磷在血液中积聚并迅速升高。肾脏病理切片结果显示病鱼肾小管上皮细胞排列松散,管腔结构不清晰,肾间质出血,伴有嗜中性白细胞浸润,这与血液磷水平的变化相吻合。本研究中发现齐口裂腹鱼嗜水气单胞菌感染后4 h血清中钙含量显著降低,随后在24 h和48 h开始逐步升高并最终与对照组差异不显著,但在攻毒后48 h血清钙含量显著高于4 h的钙含量。这可能是由于病原菌感染后对多种组织器官产生了破坏作用,机体为了应对病原菌的侵袭,需要激活多种抗菌相关酶类,从而导致机体动用更多的Ca2+参与酶促反应所致。
[1]丁瑞华.四川鱼类志[M].成都:四川科学技术出版社,1994.
[2]方 静,周 毅.齐口裂腹鱼和重口裂腹鱼消化道形态和组织结构的观察[J].四川农业大学学报,1995,13(1): 101-106.
[3]吴 青,蔡礼明,陆建平,等.齐口裂腹鱼幼鱼对水温和溶解氧的耐受力研究[J].四川畜牧兽医学院学报,2001,15(3): 20-22.
[4]若 木,王鸿泰,殷启云,等.齐口裂腹鱼人工繁殖的研究[J].淡水渔业,2001,(6): 3-5.
[5]叶 华,何丽君,程辉辉,等.注射嗜水气单胞菌灭活菌的齐口裂腹鱼外周血免疫指标的变化[J].西南大学学报(自然科学版),2014,(5): 28-35.
[6]杜宗君,汪开毓,李志琼,等.齐口裂腹鱼疾病防治初报[J].中国水产,2003,(1): 82-83.
[7]耿 毅,汪开毓,吴 麟,等.齐口裂腹鱼败血症的病原分离与鉴定[J].水利渔业,2004,(4): 71-73.
[8]杜宗君,黄小丽,汪开毓,等.齐口裂腹鱼溃疡病病原的分离鉴定[J].四川农业大学学报,2011,(2): 274-279.
[9]余泽辉,耿 毅,周 燕,等.齐口裂腹鱼无乳链球菌感染的诊断与防治[J].科学养鱼,2013,(12): 58,30.
[10]余泽辉,张 佳,耿 毅,等.齐口裂腹鱼无乳链球菌的分离鉴定及其感染的病理损伤[J].中国水产科学,2014,(6): 1244-1252.
[11]Janda J M,Abbott S L.The Genus Aeromonas: Taxonomy,pathogenicity,and infection[J].Clin Microbiol Rev,2010,23(1): 35-73.
[12]Joseph S W,Carnahan A.The isolation,identification,and systematics of the motileAeromonasspecies[J].Annu Rev Fish Dis,1994,(4): 315-343.
[13]单 红,张其中,刘强平,等.灭活菌苗免疫的南方鲇外周血液细胞免疫指标的变化[J].中国水产科学,2005,(3): 275-280.
[14]孙金辉,王庆奎,陈成勋,等.嗜水气单胞菌灭活疫苗对虹鳟免疫力和抗病力的影响[J].淡水渔业,2013,43(1): 44-49.
[15]赵 静,王 利.齐口裂腹鱼感染嗜水气单胞菌后血清指标的变化[J].水产科学,2015,(3): 178-181.
[16]Tang K F J,Lightner D V.Quantification of white spot syndrome virus DNA through a competitive polymerase chain reaction[J].Aquaculture,2000,189(1-2): 11-21.
[17]陆承平.致病性嗜水气单胞菌及其所致鱼病综述[J].水产学报,1992(3): 282-288.
[18]陈怀青,陆承平.嗜水气单胞菌:黄鳝出血性败血症的病原[J].中国人兽共患病杂志,1991,(4): 21-23.
[19]张呈念,史雨红,李明云,等.一株引起香鱼出血症的嗜水气单胞菌的鉴定[J].水产科学,2009,(7): 370-373.
[20]郭松林,关瑞章.嗜水气单胞菌感染对美洲鳗鲡不同血细胞免疫功能的影响[J].华中农业大学学报,2009,(4): 463-466.
[21]何 智,任红梅,杨德英,等.嗜水气单胞菌源黄鳝出血病的组织病理学研究[J].淡水渔业,2010,(4): 56-61.
[22]范慧慧,张呈念,安 钦,等.香鱼嗜水气单胞菌人工染病后血液生理生化指标变化[J].水产科学,2012,(5): 280-282.
[23]秦 莉,殷建国,张 薇,等.白斑狗鱼(Esoxlucius)致病性嗜水气单胞菌的分离与鉴定[J].渔业科学进展,2014,(5): 40-45.[24]Mohammadizadeh M,Afkhami M,Bastami K D,et al.Preliminary observations on the plasma composition of liza klunzingeri from the strait of Hormuz (Persiangulf)[J].Springer Plus,2013,2(1): 62.
[25]周 玉,郭文场,杨振国,等.欧洲鳗鲡“狂游病”血液生化指标研究[J].水生生物学报,2002,(3): 314-316.
[26]徐晓津,徐 斌,王 军,等.大黄鱼感染哈维氏弧菌后血液生化指标的变化及组织病理学观察[J].水产学报,2010,(4): 618-625.
[27]Tierney K B,Farrell A P,Kennedy C J.The differential leucocyte landscape of four teleosts: juvenileOncorhynchuskisutch,Clupeapallasi,CulaeainconstansandPimephalespromelas[J].J Fish Biol,2004,65(4): 906-919.
[28]Adham K,Khairalla A,Abu-Shabana M,et al.Environmental stress in lake maryut and physiological response ofTilapiazilliGerv [J].J Environ Sci Heal A,1997,32(9-10): 2585-2598.
[29]余 毅,吴宝华.白鲢疯狂病鱼的组织病理研究[J].浙江水产学院学报,1992,(2): 95-97.
[30]强 俊,杨 弘,王 辉,等.海豚链球菌感染对不同品系罗非鱼血液生化指标和肝脏HSP70 mRNA表达的影响[J].水产学报,2012,(6): 958-968.
[31]祝璟琳,柒壮林,李大宇,等.罗非鱼海豚链球菌病的病理学观察[J].水产学报,2014,(5): 722-730.
[32]储卫华.嗜水气单胞菌侵袭机制及其胞外蛋白酶的研究[D].南京:南京农业大学,2002.
[33]苗 华,潘明明.慢性肾衰竭高磷血症研究及治疗进展[J].中国血液净化,2007,(9): 500-502.
声明:本站未注明出处的转载文章是出于传递更多信息之目的。若有未注明出处或标注错误或侵犯了您的合法权益,请与本网站联系,我们将及时更正、删除,谢谢!


欢迎关注本站"养鱼第一线"微信公众帐号和养鱼第一线刘文俊视频号!将会定期向你推送本号信息!将为你精诚服务!



