

通讯:13983250545
微信:ycsh638
QQ:469764481邮箱:ycsh6318@163.com
泽蛙原蛙片虫的形态学研究
泽蛙原蛙片虫的形态学研究
邹 红 李 灿 汪润秋 李文祥 吴山功 王桂堂 李 明
(1. 中国科学院水生生物研究所, 农业部淡水养殖病害防治重点实验室, 淡水生态与生物技术国家重点实验室, 武汉 430072; 2.武汉轻工大学动物科学与营养工程学院, 武汉 430023)
蛙片虫是一类多核、无口, 渗透性营养的鞭毛类原生动物。绝大多数寄生在变温脊椎动物体内,特别是无尾两栖动物(蛙类和蟾蜍的成体及蝌蚪)的肠道内[1]。由于其皮层构造与纤毛虫颇为相似, 该类群长期以来一直被归为较原始的纤毛虫类(Protociliate)[2—4]。后来发现这类原生动物没有大小核的分化, 无性生殖为纵二分裂, 有性生殖能产生具鞭毛的配子, 故将其纳入鞭毛虫类, 并单列为蛙片亚门(Opalinata)[5,6]; 下分1纲、1目、1科, 即: 蛙片纲(Opalinatea), 蛙片目(Opalinida), 蛙片科(Opalinidae)[7]。根据蛙片虫形状和细胞核数目的不同, 可分为两个亚科, 即原蛙片虫亚科(Protoopalininae)和蛙片虫亚科(Opalininae)。
原蛙片虫属(Protoopalina)隶属于原蛙片虫亚科, 是两栖动物肠道中最常见的蛙片虫类。该属由Metcalf在1918年建立, 其典型特征是虫体常呈长棍状, 具2个或4个圆形或纺锤形胞核[3]。泽蛙原蛙片虫(P. limnocharis)是Nie(1932)首次从泽蛙肠道内发现并命名的[8]。囿于当时染色技术和观测工具的落后, 一些关键的形态特征如缝线等未能很好地予以揭示。因而, 利用现代技术手段对其形态学特征进行重新描述和修正显得尤为必要。
1 材料与方法
1.1 宿主蛙来源
泽蛙(Fejervarya limnocharis)成体210只于2016年6月采自湖北省洪湖市(北纬29°40′—29°58′N;东经113°12′—113°26′E), 活体运回并暂养于实验室。
1.2 光镜观察
活体观察将宿主蛙用100%酒精麻醉后解剖、取肠道, 置于体视显微镜Zeiss Stemi SV6/Axio Cam MRc5下观察肠道内含物。用0.65%生理盐水浸渍内含物后收集部分虫体置凹玻片上, 置于光学显微镜下观察其活体形态。
固定染色标本观察将部分虫体置胚胎皿中, 饱和升汞(HgCl2)溶液固定, 分别用埃利希苏木精(Ehrlich’s haematoxylin)和蛋白银(Protargo)染色,加拿大树胶封片, 用于观察虫体皮层及内部构造。全部观察、测量、拍照均使用德国蔡司显微成像系统Zeiss Axioplan 2 imaging and Axiophot 2完成。
1.3 扫描电镜观察
将游离出的大量虫体4000×g离心10min, 2.5%(v/v)戊二醛水溶液与2% (w/v)锇酸水溶液的1∶1混合液于4℃固定2h。用0.2 mol/L磷酸缓冲液(pH 7.4)清洗后置于涂有1%多聚赖氨酸1 cm×1 cm的玻片上。常规脱水、乙酸异戊酯置换处理后, 临界点干燥2h (HCP-2 Crtical Point Dryer)。离子溅射仪(EikoIB-3 Ion Coater)镀膜喷金后, 扫描电镜Quanta 200 (FEI, Holand)观察。
2 结果
在我们采集的210只泽蛙中, 有38只感染了泽蛙原蛙片虫。感染率为18.1%, 寄生于泽蛙直肠。固定染色标本保存于中国科学院水生生物研究所寄生虫学与协同进化学科组。
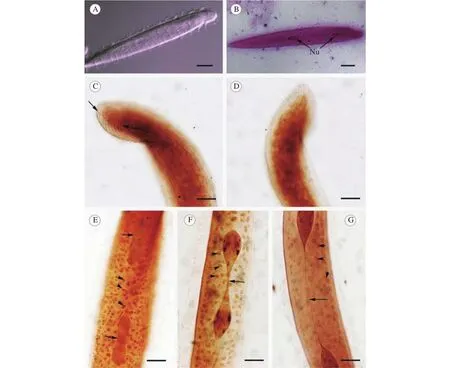
图 1 泽蛙原蛙片虫的光镜显微照片Fig. 1 Light microscope images of P. limnocharis
泽蛙原蛙片虫虫体呈长棒状, 略扁平(图 1A,B和2A)。前端凸出、稍狭窄, 约占虫体1/10 (图2A, C); 其后为虫体最宽处, 至尾端渐趋细削(图2A, D)。体长为313.8 μm (283.2—350.4 μm, n=30),体宽27.1 μm (21.6—38.4 μm, n=30)。长宽比约为10∶1。虫体周身密被鞭毛, 游动迅速且常翻转前行(图 2C)。体表皮褶形成的条纹与虫体纵轴倾斜。缝线位于虫体顶端, 长约20 μm。所有体动基列(约30—40列)均从缝线两侧发出, 沿虫体前端一直延伸至尾部(图 1C, D)。
胞核2个, 梨形; 其一在虫体前端约1/6处, 另一个在位于虫体中部略前处(图 1B)。核长19.3 μm(14.4—24.0 μm, n=30)), 宽5.5 μm (4.2—8.4 μm,n=30)。胞核分裂时从中部缢缩(图 1E), 逐渐向两端拉长并形成一条细丝(图 1F)。此细丝在虫体整个生活周期都一直存在, 成熟虫体中其长度为112.92 μm (76.8—127.2 μm, n=30)(图 1G)。另外,在胞质中可见大量大小不一、圆球状的颗粒(图1E, F和G)。
虫体相关形态学特征测量值见表 1,模式图见图 3 。
3 讨论
![]()
表 1 泽蛙原蛙片虫形态学特征测量值(单位: 微米)Tab. 1 Biometrical values (in μm) on Protoopalina limnocharis inhabiting Fejervarya limnocharis (=Rana limnocharis)
![]()
图 3 泽蛙原蛙片虫的模式图Fig. 3 Schematic drawing of P. limnocharis
泽蛙原蛙片虫是由Nie (1932)首次从泽蛙肠道内发现并命名的[8]。该蛙片虫目前仅在泽蛙中发现, 应该具有较为严格的宿主专一性。本研究中测量虫体样本所得形态特征参数值与Nie所记述值颇为相符(包括体长、体宽, 核长、核宽; 表 1)。然而,有2个重要的分类特征需要进行补充和修正: (1)缝线位于虫体顶端, 贯穿背腹侧; 所有体动基列均从缝线两侧发出, 这在前人的研究中没有描述。该结构特征与我们已经研究过的寄生于沼蛙和黑斑蛙肠道内的原蛙片虫Protoopalina pingi的缝线结构是一致的[9], 而且它们的体动基列均与虫体纵轴平行延伸, 所不同的只是两者体动基列的数目不同: P.limnocharis有30—40列, P. pingi有18—29列[9]。(2)胞核分裂时在两新核间产生的连接细丝, 在虫体整个生活周期一直存在。这一结构在其他已知的原蛙片虫属种类中未见报道, 其有何生物学功能亦不清楚。此外, 我们在观察分裂完成不久产生的P.limnocharis新生虫体时, 发现它们体内的2个胞核距离均较近且有细丝牵连; 但不同新个体中胞核的位置是有区别的: 其中一部分虫体内2个胞核均位于身体前端, 另一部分则处于虫体中部位置, 这2种位置分别对应成熟虫体中2个胞核的位置。据此推测P. limnocharis子代虫体中的2个胞核均来自于母体中同一个核, 而其两核间的连接细丝可能是胞核分裂进化历程中比较原始的残迹。
就胞核数量而言, 已知原蛙片虫的种类绝大多数为2个核, 如P. limnocharis、P. caudata michyla[10]、P. heleophrynes[11]、P. pomacantha[12]等。也有个别种类核数不定, 如P. pingi多为四核, 有时两核或六核[9]; P. quadrinucleata核数最多时可达8个[13]。作者认为这种核数不定的现象应该是在虫体分裂过程中胞质分裂和核分裂不同步造成的; 核数不定的种类应该是原蛙片虫属中进化程度较低的种类, 而核数稳定为两核的种类进化程度较高。
另外, 关于蛙片虫类的分类依据, 长期以来一直争议颇多; 但主流上基本沿用Metcalf的界定方法[4]: 双核、圆柱状种类归为Protoopalina, 双核、扁平状种类归为Zelleriella, 多核、圆柱状种类归为Cepedea, 多核、扁平状种类归为Opalina。根据我们目前已有的形态学工作基础(特别是银染标本)和分子生物学数据(全rDNA序列), 我们认为这种分类方法是基本正确的。这将在我们的研究中进一步补充、求证并最终得到较为合理的结论。
参考文献:
[1]Mignot J P. Patterning in opalinids. I. Implications of new morphological and ultrastructural findings on the genesis of kinetics [J]. European Journal of Protistology, 1994,30(2): 196—210
[2]Boisson C. Deux nouvelles opalines de l’Ouest africain[J]. Bulletin de i’Institut Français d’Afrique Noire, 1965,27A: 858—865
[3]Metcalf M M. Opalina and the origin of the Ciliata [J].Anatomical Record, 1918, 14: 88—89
[4]Metcalf M M. The opalinid ciliate infusorians [J]. Bulletin of the United States National Museum, 1923, 120:1—484
[5]Corliss J O. The opalinid infusorians: flagellates or ciliates [J]? Journal of Eukaryotic Microbiology, 1955,2: 107—114
[6]Corliss J O. Flagellates, opalinids and the search for the most primitive ciliate and its progenitor [J]. Ceylon Journal of Science, 1979, 13: 65—78
[7]Sheng Y F, Wang J G. Protozoology [M]. Beijing: Science Press. 1999, 223—231 [沈韫芬, 汪建国. 原生动物学. 北京:科学出版社. 1999, 223—231]
[8]Nie D S. On some intestinal ciliates from Rana limnocharis Gravenhorst [J]. Zoological Series, 1932, 6:183—199
[9]Li W, Wang C, Huang F, et al. Redescription of Protoopalina pingi Nie, 1935 inhabiting the recta of Hylarana guentheri and Pelophylax nigromaculatus in China [J].Parasite, 2014, 21: 46
[10]Nie D S. Intestinal ciliates of amphibian of Nanking [J].Contributions from the Biological Laboratory of the Science Society of China: Zoological Series, 1935, 11:67—95
[11]Delvinquier B J, Markus M B, Passmore N I. Opalinidae in African Anura. Genus Protoopalina [J]. Systematic Parasitology, 1995, 30: 81—120
[12]Grim J N, Pérez-España H, Martínez-Díaz S F. The morphology of Protoopalina pomacantha, n. sp., symbiont in the rectum of the Angelfishes, Pomacanthus zonipectus and Holacanthus passer. A light, scanning electron and transmission electron microscopic study [J]. European Journal of Protistology, 2000, 36(3): 343—350
[13]Lu K. On some parasitic ciliates from frogs of Pehpei [J].Sinensia, 1945, 16: 65—72
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!



