本站搜索
西南渔业网 水花鱼 水产养殖专业网 水产前线网 西南水产网 民间水产网 丰祥渔业网 永川水花网为您精诚服务!

最新头条
- 我国原产地观赏鱼及其资源保护与发展
- 大苏打在水产养殖中的应用技术
- 加州鲈帆布池养殖效益高
- 长江刀鲚人工繁育技术研究进展
- 贝壳的结构与应用的研究现状
- 硅藻休眠孢子的生活史与研究意义
- 不同年龄细鳞鱼的繁育效果
- 塑料盒养甲鱼,2个月亏了近百万元!水产养
- 阿司匹林再“立功”!哈佛医学院最新:10
- 农药稀释倍数和用水量如何转换?用好一个公
- 淘宝,“仅退款”突然有调整!
- 金龙鱼国际商业贸易规则与实践
- 浅谈水生野生动物保护管理
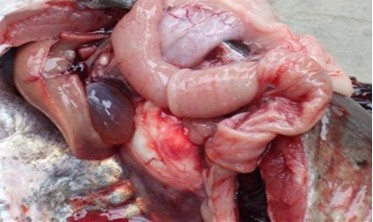
- 大鲵腹水病解剖观察与防治实例
- 饥饿对家养孔雀鱼色彩的影响
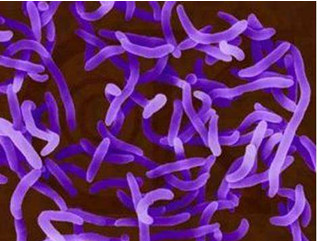
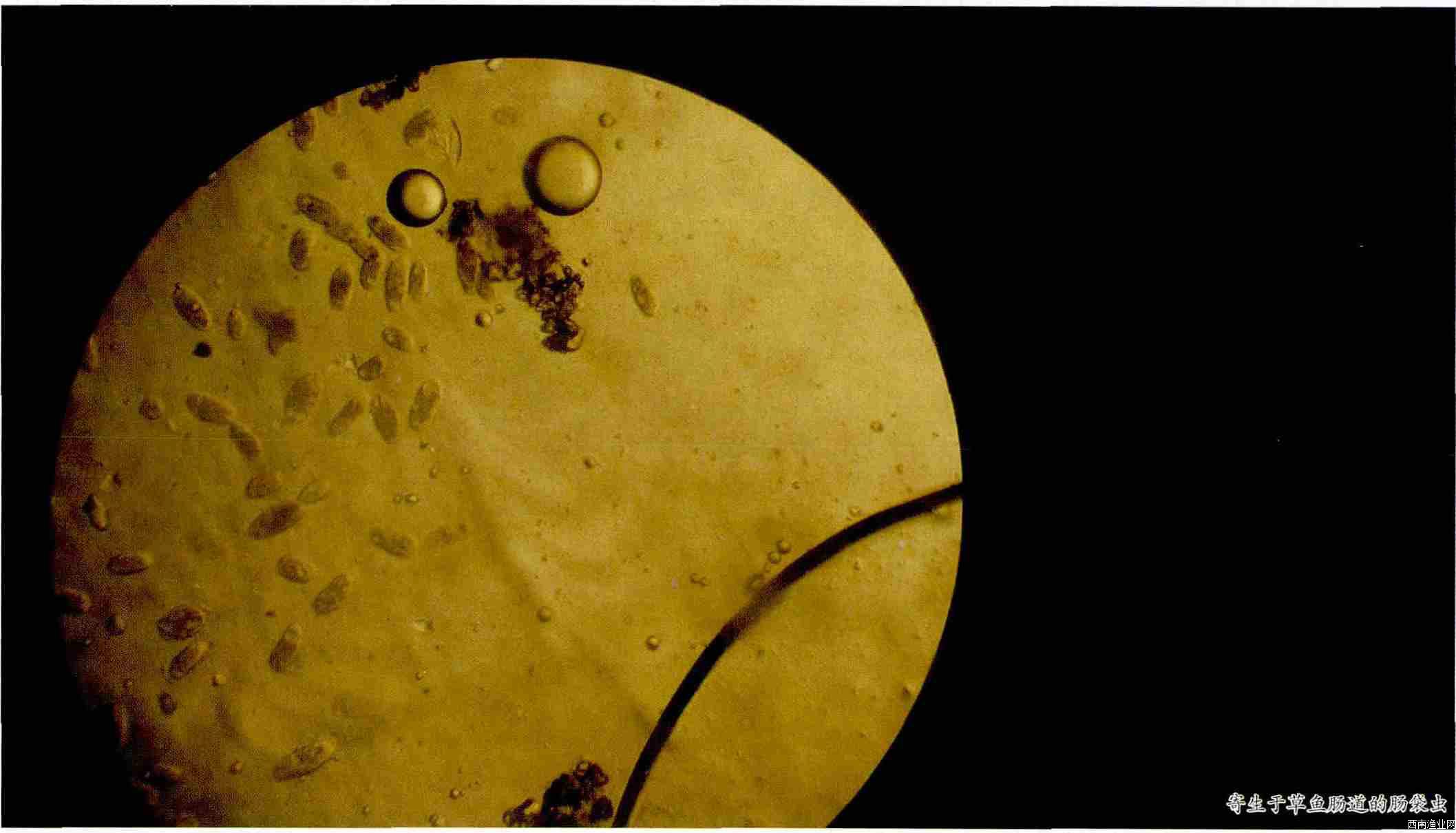
- 浅析淡水鱼养殖发病原因及防治对策
- 水族馆展示设计的两个创意
- 中国渔业增殖放流问题及对策研究
- 鱼籽营养成分及相关产品的研究进展
- 遭遇假水花,苗场亏损达30万,多次交涉无


热门内容